
病毒引起的线粒体生物能学变化机制可能成为一种治疗传染病的潜在方案
- 北京华威中仪科技有限公司2022年11月1日 2:52 点击:426

由病毒所引起的传染性疾病致死率占全世界人口死亡的很大比例。传染病的种类和特点多种多样,在全球范围内,多种传染病病毒可对非特定环境和人群进行大范围的传染,例如,革登热等病毒的感染不因环境改变而限制,它可造成大范围的流行性传播。如果考虑到与传染病病原具有显著相关的慢性疾病,例如肝细胞癌(乙型肝炎或丙型肝炎病毒等),那么传染病的覆盖度和死亡率将会更高。
宿主与病毒之间的相互作用包括两种机制,即宿主代谢在病毒复制中的重要作用和线粒体在此过程中的主动参与。病毒没有进行能量代谢的生理机制,因此他们biyu寄生生活在宿主细胞内才能完成繁殖与传播。各种病毒会应用其自身独特的生理机制来调节线粒体的产能,以便于提高病毒的自我复制能力。在近期的研究中,发现了宿主与病毒相互作用的一个新观点,即宿主代谢在病毒复制和感染过程中起到了关键性作用。
病毒的繁衍循环是一个动态过程,涉及多种不同的宿主细胞器。不同的病毒已经演化出其独特的繁衍策略。实际上,线粒体除了在细胞凋亡过程中发挥作用外,也积极的参与了病毒的复制过程。以下将详细阐述在病毒感染过程中线粒体生物能的调节模式以及病毒诱导的线粒体功能障碍对细胞代谢的影响。
1、细胞器功能:线粒体ATP合成
少量的ATP是通过真核细胞胞质中的糖酵解合成的,绝大多数细胞ATP是由膜结合蛋白复合物合成的。跨线粒体内膜的pH梯度和跨膜电势的联合作用所产生的电化学能称为质子原动力,当质子通过蛋白复合物返回线粒体基质时,它驱动ADP和Pi合成ATP,利用的是复合蛋白物 V(ATP合酶)通道。线粒体很容易受损,线粒体一旦受损则会对细胞功能造成严重后果。例如,线粒体膜通透性(MMP)的病理状况会影响氧化磷酸化和线粒体ATP合成。完整细胞或通透性细胞中测得的呼吸控制比和P / O比可用于评估不同病理状态中的线粒体功能是否完整 ,包括病毒感染的宿主细胞线粒体功能情况。这些测量结果可用于评估磷酸化能力和电子传递能力、呼吸复合物的活性以及偶联效率等,是研究病毒对线粒体功能的影响以及了解病毒性疾病发病机理的重要工具。
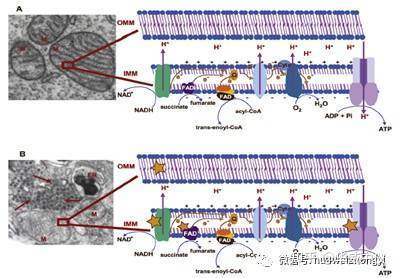
图1.线粒体呼吸链和氧化磷酸化:(a)人肝细胞的电子透射显微照片,显示线粒体(M)带有电子致密cr。(b)感染登革热病毒的人类肝细胞的电子传输显微照片(电子致密颗粒;箭头)。病毒颗粒(红色箭头)位于线粒体(M)和内质网(ER)附近。[YL1]
2、细胞器病理:病毒感染期间线粒体的生物能
抗病毒感染后产生的先天免疫反应主要以I型干扰素(IFN-α和IFN-β)、其他促炎性细胞因子和趋化因子为主。感染后,宿主的种系编码模式识别受体(PRR)可以检测到病毒成分。胞质维甲酸诱导基因I(RIG-1)样受体的信号转导取决于线粒体抗病毒信号蛋白(MAVS)(也称为IPS-1,CARDIF和VISA)。MAVS与线粒体外膜(OMM)相关联,其定位对于抗病毒免疫反应至关重要。MAVS抗病毒信号传递的一个显着特点是它依赖于线粒体的动力学。调节线粒体融合的线粒体融合蛋白与抗病毒信号蛋白(MAVS)相互作用,这对于正确的抗病毒反应至关重要 。线粒体网络的完整性和持续的呼吸能力对于早期抗病毒的先天免疫反应很重要。
线粒体控制病毒感染过程中的凋亡也是先天免疫的重要方面。抗凋亡或促凋亡的病毒蛋白共同选择宿主线粒体来调节MMP,从而影响线粒体生物能,进而有利于病毒复制 。病毒诱导的线粒体生物能学变化是驱动复制的重要机制,代表了对病毒感染的早期反应。代谢组学和通量组学分析也提示了人类巨细胞病毒HCMV感染期间改变了线粒体的功能,增加了糖酵解和TCA循环,使代谢通量增加 。
如:登革热病毒感染是z常见的虫媒病毒疾病,会引起急性感染,进而引起生物体的肝功能障碍;Sindbis病毒(典型的甲型病毒)在小鼠中可引起急性脑炎,它是研究人类脑炎的分子机制的重要模型。线粒体生物能学改变可能参与了这一过程;在慢性丙型肝炎病毒(HCV)感染中也观察到线粒体功能的严重改变,这是肝病致死的主要原因。稳定表达HCV多蛋白的人细胞表现出常规呼吸减少和与ATP合成无关的呼吸减少,从而导致RCR降低。
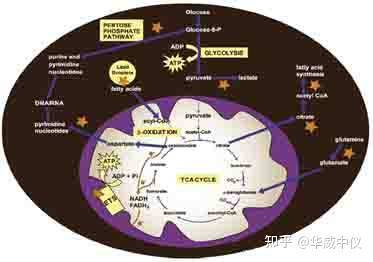
图2.病毒感染的代谢作用:病毒复制影响线粒体生物能,进而影响整体能量代谢。星形和粗箭头表示受病毒感染刺激的代谢步骤和治疗中潜在的代谢靶标,如文中所述。在HCMV,Mayaro和Kaposi-肉瘤病毒感染后,已观察到通过糖酵解和戊糖磷酸途径的通量增加。单纯疱疹病毒1通过使草酰乙酸偏离TCA周期而刺激嘧啶的代谢;柠檬酸盐偏离线粒体用于脂肪酸合成是HCMV利用的机制;登革热病毒刺激脂质液滴储存的降解并增加_氧化;另一方面,HCV感染会导致脂质滴中三酰基甘油的利用率降低;HCMV感染后,观察到谷氨酰胺代谢使碳进入TCA循环的增加;病毒感染后,线粒体电子转移系统(ETS)受到不同方式的影响。
实验证明在接受抗病毒治疗的患者中,从感染人类免疫缺陷病毒(HIV)的患者中分离出的CD4 +淋巴细胞的耗氧量显着降低,ETS和氧化磷酸化能力均明显降低。抗逆转录病毒治疗本身会引起这种效应。因为未接受治疗的患者的淋巴细胞线粒体呼吸与对照组相似。因此,线粒体生物能学的调节是病毒诱导的细胞代谢变化的重要方面,了解这些作用可能有助于选择适当的抗病毒治疗方法和后续治疗工具。
3、细胞生理学:病毒引起的线粒体功能障碍(对细胞代谢的影响)
细胞具有ATP调节和ATP利用的生理机制,因此细胞ATP含量保持恒定。癌细胞中线粒体和细胞质之间的交换发生着变化,即使在较高氧的分压下,也会通过糖酵解的增加确保 ATP的产生速率。例如,在卡波西氏肉瘤相关疱疹病毒的潜伏感染中,线粒体耗氧量的减少可通过糖酵解的增加来补偿 。糖酵解抑制剂如草酸盐和2-脱氧葡萄糖可诱导感染细胞的凋亡。在长期表达HCV多蛋白的细胞模型中也观察到了糖酵解补偿,该蛋白表现出抑制的线粒体功能。糖酵解的增加是由HIF-1稳定介导的,它不仅维持而且还增加了细胞ATP的含量 。在HCM 和单纯疱疹1病毒HSV-1感染之后,同样观察到细胞ATP的增加。正如我们在Mayaro感染中观察到的那样,即使在没有线粒体改变的情况下,糖酵解的上调也被观察到,这也是由α病毒引起的。葡萄糖代谢会随着PFK-1活性的增加而改变,这不仅会驱动ATP的合成,而且还会使戊糖磷酸途径(PPP)的葡萄糖碳偏向生物合成目的。在HCMV感染的细胞中也发现通过PPP的通量增加(图2)。HCMV感染后也显示了PFK-1的功能激活。
病毒感染还可通过调节TCA周期影响脂质代谢。HCMV就是这种情况。HCMV感染将柠檬酸从线粒体转移到细胞质,在那里它充当ATP-柠檬酸裂解酶的底物,产生草酰乙酸和乙酰辅酶A。后者用作脂肪酸合酶复合物的间接底物以合成脂肪酸(图2)。通过抑制脂肪酸的合成,研究者观察到了较低的病毒产生,表明脂质生物合成的上调对于病毒的包膜和复制很重要。除了代谢中间体的偏离外,固醇调节元件结合蛋白1(SREBP1)的活化是脂肪生成上调和HCMV增殖的关键因素。在不同的HCV感染模型中也观察到脂质代谢的惊人改变。HCV感染的细胞(以及感染对象的肝脏活检)显示出三酰基甘油的积累增加,并沉积在LD中。
在单纯疱疹病毒1(HSV-1)感染期间也观察到TCA周期的调节。通量组学分析表明,感染HSV-1的细胞偏离了TCA周期的草酰乙酸,并增加了其转化为天冬氨酸的嘌呤嘧啶的合成(图2)。这是一个非常有趣的发现,显示了另一种依赖中间代谢的病毒复制策略,如在HCMV感染中观察到的。
此外,由于不同的病毒能够调节抗氧化酶,病毒感染会诱导控制复制的活性氧(ROS)的产生。ROS产生的增加可能有助于线粒体生物能学的改变。反过来,功能异常的线粒体可加剧氧化应激,导致能量崩溃和细胞死亡。钙代谢与ROS诱导的线粒体氧化应激有关,如HCV感染,内质网(ER)积极参与这一过程。z近,由于钙调蛋白依赖性激酶抑制作用降低了糖酵解通量并阻止了病毒复制,因此钙代谢也被证明参与了HCMV诱导的糖酵解,提示了另一种可能的治疗靶点。病毒诱导的ROS产生也将免疫应答与线粒体联系起来。ROS产生增加会增强MAVS向IRF-3和NF_B的下游信号传导,因此限制了病毒复制。线粒体(通过巨噬自噬选择性去除线粒体)还通过去除功能异常的线粒体来减弱ROS的产生,这是控制加剧免疫反应的一种方法。需要确立线粒体作为控制不同病毒复制机制的影响。
病毒感染会引起新陈代谢的重新编程,从而导致独特的生物能表型,这对于驱动病毒复制至关重要。因此,代谢拮抗剂不仅对治疗病毒感染很重要,而且对我们更好地了解宿主代谢的病毒调节(尤其是不同病毒使用的代谢策略)也很重要。
关于线粒体超复合体的组织和感染细胞中线粒体网络的功能和结构研究是线粒体生理的重要方面。这些方面都可能阐明了病毒策略,因为它们都与钙代谢和氧化应激的调节有关。
参考文献:https://www.ncbi.nlm.nih.gov/pubmed term=Virus induced%20changes%20in%20mitochondrial%20bioenergetics%20as%20potential%20targets%20for%20therapy
--------------------------------------------------------------------------------------
奥地利OROBOROS O2k光电联合多维度能量代谢检测整体解决方案
 奥地利OROBOROS O2k细胞能量代谢分析仪
奥地利OROBOROS O2k细胞能量代谢分析仪
1、光电联合多参数实时动态检测技术:
电化学实时动态检测模块:pO2(高分辨率极谱氧电极传感器,耗氧率检测分辨率为±1 pmol O₂∙s-¹∙mL-¹)、pH、H2O2、TPP+(测量线粒体膜电位)、H2S、NO、质体醌;荧光实时动态检测模块:MMP(测量线粒体膜电位)、ATP、Ca2+、ROS、NADH;
2、拥有原代细胞、原代组织能量代谢快速检测技术:
无需过夜培养、样品快速检测,更加客观反应样品更接近体内的能量代谢水平;
3、拥有多维度能量代谢分析平台:
适应线粒体、细胞、组织块、活检样品等不同层次不同水平的样本检测。
关 于 公 司
北京华威中仪科技有限公司成立于2006年,历经多年的积累与沉淀,华威公司已经成为国内经多识广的实验室设备的供应商/技术服务商。
北京华威公司深耕中国细胞能量代谢市场十二年,为全国的科研用户提供从组织-细胞-线粒体一站式能量代谢解决方案。业务涵盖细胞能量代谢技术交流/实验技术支持/DEMO技术展示/设备销售/应用技术指导等。
生物体能量代谢研究适用于多种学科:运动生理学、线粒体医学、老龄化医学、干细胞医学、癌症、肥胖、糖尿病、氧化应激、药理与毒理、心血管类疾病、神经退行性病变等。
生物体能量代谢在科学研究中具有重要的论证地位,细胞器线粒体作为能量供应的主要场所,参与生物体内的绝大多数代谢过程,多种疾病的病理成因与线粒体损伤有关,通过预防线粒体损伤的发生、进行线粒体的移植治疗以及促进线粒体的再生与自噬,往往能够靶向针对疾病的病灶位点形成快速有效的治疗策略,从而达到早预防早治疗的目的;同样,能量代谢为生物起源、线粒体发生以及生态适应性等多个领域提供生物进化的直接或间接证据,也为研究多样化生态环境适应性、ji端环境能量保障、能量供应大数据等方面提供理论基础。
华威公司全国区代理产品
德国INCYTON实时全息多参数细胞能量代谢分析系统
奥地利OROBOROS O2k细胞能量代谢分析仪
积极乐观、科学创新是我们华威的精神,在过去十六年的工作中得到了广大用户的认可和高度评价,我们也会一如既往的坚持优良产品、更好的服务继续服务于中国的科研事业。
我们还可以为您提供其他的能量代谢产品
 德国INCYTON实时全息多参数细胞能量代谢分析系统
德国INCYTON实时全息多参数细胞能量代谢分析系统
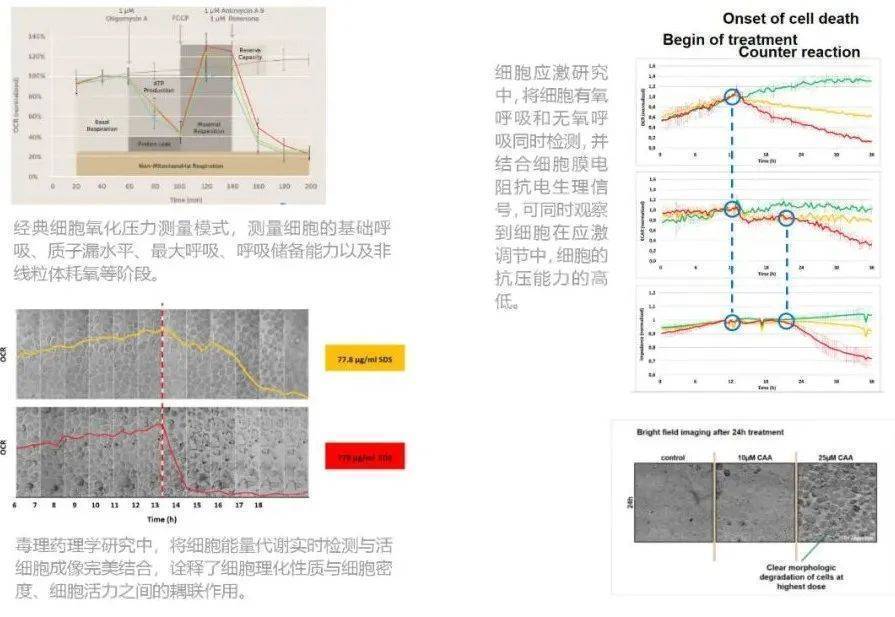
Ø 24孔板每孔单独进行实验,独立切向流换液、加药系统;
Ø 7个试剂池,可实现6种药物重复、多次准确加入及更换;
Ø 同时检测产酸率、耗氧率、细胞膜电阻抗;
Ø 实现实时影像跟监测相结合的仪器;
Ø 全自动移液站,可实现连续监测几小时、几天、几周至数月的长期实验。
联系邮箱:kefu@labbase.net
版权与免责声明
- 凡本网注明“来源:来宝网”的所有作品,版权均属于来宝网,转载请必须注明来宝网, //www.next-search.com,违反者本网将追究相关法律责任。
- 本网转载并注明自其它来源的作品,目的在于传递更多信息,并不代表本网赞同其观点或证实其内容的真实性,不承担此类作品侵权行为的直接责任及连带责任。其他媒体、网站或个人从本网转载时,必须保留本网注明的作品来源,并自负版权等法律责任。
- 如涉及作品内容、版权等问题,请在作品发表之日起一周内与本网联系,否则视为放弃相关权利。





